

出品:科普中国
作者:吕泽龙(中国科学院动物研究所)
监制:中国科普博览
提起鸟类,你脑海中会浮现怎样的画面?是展翅高飞的雄鹰,还是城市公园中随处可见的麻雀?还有些了解古生物的人会立刻联想到——恐龙。没错,基于化石材料和系统分类学,鸟类正是恐龙总目(Dinosauria)、蜥臀目、兽脚亚目的分支。并且,一些兽脚亚目恐龙也和鸟一样长有羽毛。
不过,与现代鸟类的羽毛相比,这些恐龙的羽毛形态更加多样:有些类似头发、鸡鸭的绒羽,有些则是片状对称或不对称羽毛。例如,科学家们基于雷神翼龙(Tupandactylus)等翼龙的化石研究,发现翼龙体表就覆有丝绒状的羽毛。
这些羽毛究竟从何而来?在鸟类、非鸟类恐龙以及翼龙身上,羽毛可能扮演着多重角色——或许是飞行的工具,也许是求偶时的华丽装饰,又或者是作为伪装的外衣。本篇文章,让我们一同深入探索其中的奥秘。
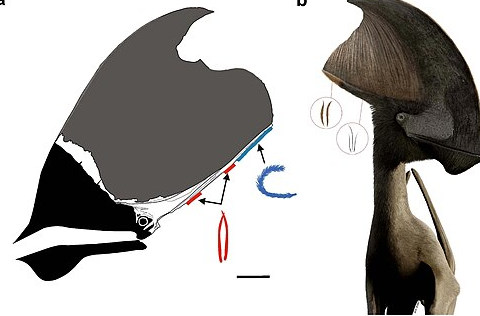
雷神翼龙及其原始羽毛
(图片来源:维基百科)

兽脚亚目恐龙羽王龙(Yutyrannus)的尾部化石,保留有羽毛证据
(图片来源:维基百科)
羽毛形态的演变之路
其实,羽毛最初的形态并非我们所熟知的那样,而是有点类似于丝绒或者头发。不过它们的组成成分与人类头发并不一样,人类头发主要成分是阿尔法角蛋白,而它们的成分里面主要是贝塔角蛋白。
早期的羽毛是从皮肤表面延伸出来的丝状结构,呈中空圆柱形且没有分叉,可能用于伪装或者装饰,也有人认为这样的功能可以维持体温。之后,这些羽毛的结构逐渐改变——分叉且呈现丝状。再之后出现了羽轴,又形成对称的片状羽毛。

具有片状羽毛的赫氏近鸟龙标本
(图片来源:作者拍摄于国家自然博物馆)
圆柱形且中空的丝状结构羽毛在天宇龙、鹦鹉嘴龙、古林达奔龙身上发现过,这些恐龙并非蜥臀目的兽脚类恐龙,而是属于鸟臀目恐龙。这表明,这些原始的羽毛可能起源于鸟臀目恐龙、蜥臀目恐龙和翼龙的最近共同祖先,只不过在后续演化道路上,有些成员失去了羽毛,比如体型庞大的三角龙和真蜥脚类恐龙。而另一些类群则进一步强化了羽毛的发育,如上文提到的雷神翼龙,就有丝状不分叉羽毛与丝状分叉羽毛两种。

原始中华龙鸟化石模型,有原始的丝状羽毛
(图片来源:作者拍摄于国家自然博物馆)

天宇龙复原图
(图片来源dinosaurpictures.org)
在兽脚类恐龙的廓羽盗龙类(Pennaraptora)分支中(包括窃蛋龙类、鸟翼类、驰龙科和伤齿龙科等),演化出了对称片状羽毛,并带有“羽轴”——羽毛的“脊梁骨”。虽说这些羽毛的空气动力学性能仍然有限,但是可以在求偶时用来展示。廓羽盗龙类的胡氏耀龙(Epidexipteryx hui)就是典型的例子,学界认为其尾巴上明显的修长尾羽可能就是装饰,用以吸引异性。

珍珠鸡羽毛,红色圈内较粗的“分割线”为羽轴
(图片来源:维基百科)

胡氏耀龙标本,有尾巴羽毛
(图片来源:维基百科)
当片状羽毛演化出不对称结构时,往往预示着飞行能力的出现。仔细观察这些片状羽毛的两侧,会发现羽毛的侧面有很多“毛”,这些类似枝条的“毛”称之为羽枝。羽枝又是由带着沟槽的羽小枝构成的,羽小枝盘根错节地排列,依靠它们的羽小钩互相“咬”在一起形成沟槽连锁机制。羽小枝排在一起的同时,这些“锁”不能在受到外力时散开,为此,羽小枝上还带有结节结构来增强固定。为什么要固定上呢?因为只有固定起来才可以让用于飞行的羽毛足够“强韧”,抗撕裂。
抗撕裂的飞行羽毛并非所有恐龙的共同特征。在廓羽盗龙类中,伤齿龙科(Troodontidae)和驰龙科(Dromaeosauridae)这些近鸟类非鸟恐龙虽然拥有片状羽毛,但它们羽毛片中间的羽轴相对纤细,而且羽小枝排列很松散,没有强韧的连锁结构。
反鸟类属于鸟类但是并非现代鸟类,它们则拥有变厚的羽轴这一新特征,但是羽小枝之间排列依旧不够紧密,相邻的羽小枝之间存在空隙,这就导致它们飞行空气动力学效率低。相比之下,这些活到今天的恐龙后裔——现存的鸟类,其羽毛能抗撕裂,羽小枝紧密排列在一起,飞行效率显著较高。

反鸟类的代表类群之一——渤海鸟类
(图片来源:作者拍摄于国家自然博物馆)

顾氏小盗龙化石
(图片来源:作者拍摄于国家自然博物馆)
近期研究中,科学家发现始祖鸟具有三级飞羽,而很多类似鸟的非鸟类恐龙则没有这个结构。当羽小枝排列较为松散时,就会形成鸟类和部分非鸟类恐龙拥有的绒羽。鸟类是廓羽盗龙类下面鸟翼类(Avialae)的分支,从晚侏罗世到晚白垩世期间,其骨骼结构经历了显著变化,物种数目也有所增加。鸟翼类中的真鸟类分支包括现代鸟类,其胸骨等结构和反鸟类存在差异。
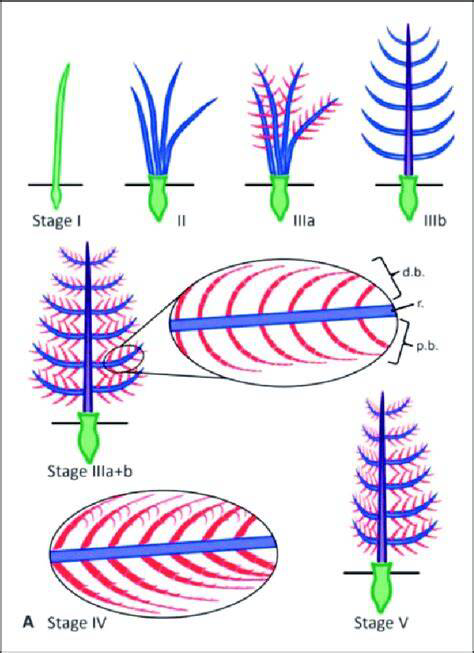
从丝状纤维(Stage Ⅰ)到分叉(II),再到羽轴和羽小枝形成(IIIa和IIIb),最后羽小枝排列起来(Stage IV),不对称飞羽形成(Stage V)
(图片来源:参考文献[15])
羽毛是如何发育形成的?
为了找出羽毛发育形成的奥秘,科学家们用驯化红原鸡(也就是家鸡)的胚胎进行了实验,尝试让鸡全身长出丝状的原始羽毛。
如何诱导原始羽毛的生长呢?关键在于SHH信号通路,该通路能影响鸟类体表羽毛和裸区鳞片的发育。2023年就有科学家通过促进这一通路,成功使鸡腿部原本光滑的鳞片转变为羽毛。
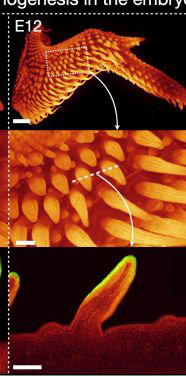
鸡胚胎12天的原始羽毛
(图片来源:参考文献[3])

鸡爪的鳞片转变为羽毛实验
(图片来源:参考文献[17])
考虑到鸡胚胎在发育的过程中,羽毛原基会发育成相应的羽毛,因此研究人员最初选择在胚胎发育第9天(羽毛原基尚未长成),开始用药物抑制这一基因信号通路。然而,实验并未达到预期效果——虽然药物在早期阶段显示出抑制作用,但从鸡胚胎第14天起,鸡羽毛逐渐发育为结构复杂的绒羽和片状羽毛等。
这一结果表明,从龙到鸟类的羽毛性状结构变得复杂,依赖的是复杂的基因调控网络的共同作用,调控网络也可以在环境干扰下保证羽毛发育。当然,在实验中那些被加入药物抑制基因信号通路的鸡胚胎,在孵化成为小鸡之后,比起未加入药物的对照组小鸡,身上具有更多的“裸区”没有羽毛。虽说这些小鸡在成长的过程中,身上的“裸区”也长出了结构复杂的羽毛。
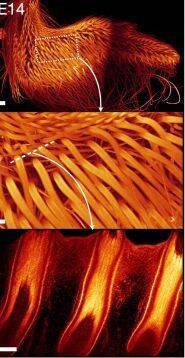
14天的鸡羽毛
(图片来源:参考文献[3])
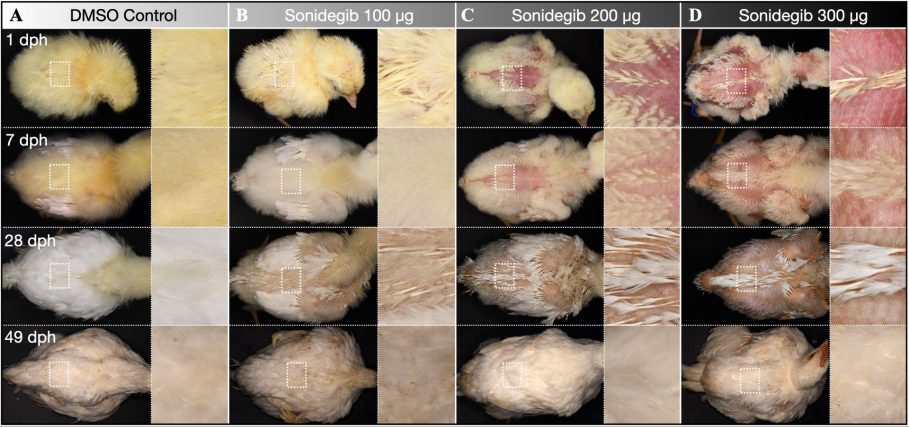
胚胎分别加入不同量抑制药物的小鸡的羽毛生长情况(从左到右依次是对照组,100微克,200微克和300微克),后面三个在孵化之初,体表有明显较多裸区
(图片来源:参考文献[3])
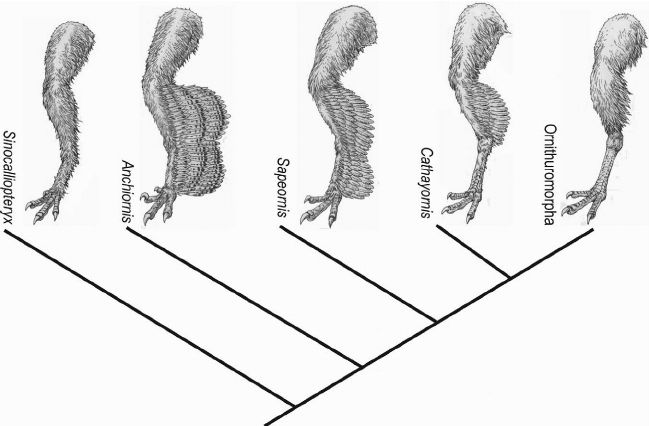
从左到右依次是并非鸟翼类的中国龙鸟,鸟翼类原始的近鸟龙,鸟翼类的会鸟,反鸟类的华夏鸟和现代鸟类所属分支,可以看出鸟翼类腿上的毛减少了
(图片来源:参考文献[14])
当鸟类演化出羽毛时,其体表的鳞片也并非全都消失了。基于对鸟翼类恐龙(包括鸟类)腿部羽毛的分析发现,原始的鸟翼类恐龙在朝着鸟类演化过程中,退化了腿部羽毛,重新发育出鳞片。这一演化特征在现代家鸡的足部鳞片中得到了典型体现。
羽毛的特殊功能
在功能上,科学家们还发现了不同鸟类和恐龙羽毛的特别用处。除了我们熟悉的蓝孔雀、红原鸡等鸟类的性展示功能,鸟身上还有纤羽(hair feather)。这类羽毛可以作为感受器,主要功能是感知正羽(上文提到的片状的羽毛,有对称和不对称两种)的姿态。

纤羽
(图片来源:allaboutbirds)

位于尾部对称的正羽
(图片来源:allaboutbirds)
在相关肌肉(直立肌和抑制肌,前者负责立起来羽毛,后者反之)的控制下,羽毛可以进行“伸缩”,当羽毛被提起来的时候,鸟类就会呈现“炸毛”状态。一些鸟类头上的羽毛就能以此表示它们受到了惊吓。比如凤头鹰(一种常见于中国南方的国家二级保护动物)、红角鸮(我们俗称“猫头鹰”的一种)就是典型代表,在受惊时会立即竖起头上的羽毛。

没有立起头冠的凤头鹰
(图片来源:作者拍摄于国家动物博物馆)
不过,鸟儿“炸毛”除了受到惊吓,还有其它情况。比如有些是为了散热,竖立羽毛,让平时没有接触到外界空气的皮肤不被闷着。还有的鸟类是为了“伪装”,就像南美洲的栗斑翅伞鸟(Laniocera hypopyrra)会在幼年期晃动自己的羽毛,假装自己是有毒的绒蛾科(Megalopygidae)毛毛虫。

热天下让羽毛动起来的家燕
(图片来源:作者拍摄于北京奥森公园)

栗斑翅伞鸟和它伪装出的虫子
(图片来源:sci)
羽毛的适应性转变
对于不会飞的鸟而言,其羽毛的功能发生了显著的适应性转变。以鸮鹦鹉和渡渡鸟为例,它们翅膀短小,腿部强健,但保留了不对称的片状羽毛,且初级飞羽的数目依旧是9-11根。相比之下,南方鹤鸵、大美洲鸵和小斑几维等鸟类的羽毛则呈现出更为明显的退化特征:不仅丧失了飞行功能,其羽片结构也趋于简化(如鹤鸵的毛发状羽毛与布偶猫的被毛相似),初级飞羽数目也有所改变。

大美洲鸵,羽毛看着更像毛
(图片来源:作者拍摄于上海动物园)
这背后的原因又是什么呢?学界对30类不飞行鸟类及其近缘物种的骨骼和羽毛进行对比发现,在飞行能力退化过程中,这些鸟类的腿部和翅膀演化速率更快,而羽毛形态演化速率相对慢。这是因为腿部和翅膀长短对于地面生活更为重要,而羽毛生长需要的能量相对骨骼与肌肉发育更低,因此演化相对滞后。羽毛的演化主要表现为不对称性逐渐降低,甚至变得像小斑几维那样只剩下绒毛。同时,翅膀上的初级飞羽数量也不再保持飞行鸟类典型的9-11根,而是出现不同程度的增减。
此外,羽毛性状的改变还与飞行能力丧失的时间节点有关。在漫长的演化历程中,鸟类首先从有齿、长尾的鸟翼类恐龙演化为具备飞行能力的现代鸟类,随后部分类群又再度放弃或削弱了飞行本领(最早可追溯至白垩纪晚期)。其中,比起南方鹤鸵、大美洲鸵和企鹅等很早丧失飞行能力鸟类,较晚丧失飞行能力的鸟类羽毛“爆改”程度更低。
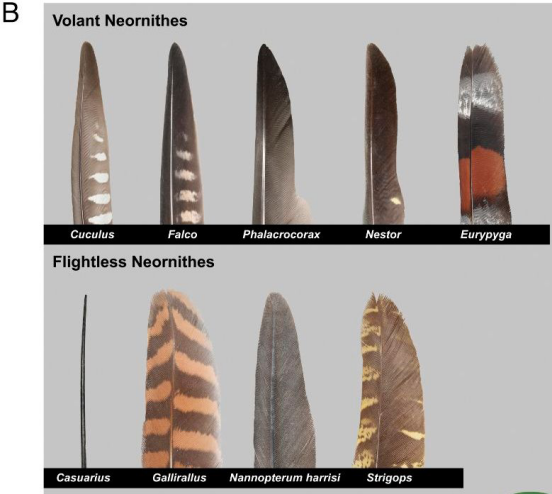
上图为飞行鸟类不对称飞羽
下图为不飞鸟类的羽毛示例,其中有些留有不对称飞羽,但是有些羽毛类似头发,如最左侧的鹤鸵羽毛。
(图片来源:参考文献[12])
由此观之,从恐龙到现代鸟类,羽毛在演化过程中展现出惊人的可塑性。它不仅能够适应飞行需求,在丧失飞行能力的类群中还表现出多样化的形态与功能转变,充分体现了其对不同生态环境的卓越适应能力。
参考文献:
[1]Lingham-Soliar T, Murugan N. A new helical crossed-fibre structure of β-keratin in flight feathers and its biomechanical implications. PLoS One. 2013;8(6):e65849.
[2]Cincotta A, Nicolaï M, Campos HBN, et al. Pterosaur melanosomes support signalling functions for early feathers. Nature. 2022;604(7907):684-688.
[3]Cooper RL, Milinkovitch MC. In vivo sonic hedgehog pathway antagonism temporarily results in ancestral proto-feather-like structures in the chicken. PLoS Biol. 2025;23(3):e3003061.
[4]O'Connor J, Clark A, Kuo PC, et al. Chicago Archaeopteryx informs on the early evolution of the avian bauplan. Nature. Published online May 14, 2025.
[5]Kiat Y, O'Connor JK. Functional constraints on the number and shape of flight feathers. Proc Natl Acad Sci U S A. 2024;121(8):e2306639121.
[6]Pan Y, Zheng W, Moyer AE, et al. Molecular evidence of keratin and melanosomes in feathers of the Early Cretaceous bird Eoconfuciusornis. Proc Natl Acad Sci U S A. 2016;113(49):E7900-E7907.
[7]Pan Y, Zheng W, Sawyer RH, et al. The molecular evolution of feathers with direct evidence from fossils. Proc Natl Acad Sci U S A. 2019;116(8):3018-3023.
[8]Padian K. 25th anniversary of the first known feathered dinosaurs. Nature. 2023;613(7943):251-252.
[9]Saitta ET, Balaji L, Mitchell JS, Makovicky PJ. Feather evolution following flight loss in crown group birds: relaxed selection and developmental constraints. Evolution. 2025;79(5):737-751.
[10]Homberger DG, de Silva KN. The role of mechanical forces on the patterning of the avian feather-bearing skin: A biomechanical analysis of the integumentary musculature in birds. J Exp Zool B Mol Dev Evol. 2003;298(1):123-139.
[11]Saitta ET, Balaji L, Mitchell JS, Makovicky PJ. Feather evolution following flight loss in crown group birds: relaxed selection and developmental constraints. Evolution. 2025;79(5):737-751.
[12]Kiat Y, O'Connor JK. Functional constraints on the number and shape of flight feathers. Proc Natl Acad Sci U S A. 2024;121(8):e2306639121.
[13]Xu X, Currie P, Pittman M, et al. Mosaic evolution in an asymmetrically feathered troodontid dinosaur with transitional features. Nat Commun. 2017;8:14972.
[14]Zheng X, Zhou Z, Wang X, et al. Hind wings in Basal birds and the evolution of leg feathers. Science. 2013;339(6125):1309-1312.
[15]Farzeen Shahid, Jing-Shan Zhao, Pascal Godefroit. Design of flying robots inspired by the evolution of avian flight. ARCHIVE Proceedings of the Institution of Mechanical Engineers Part C Journal of Mechanical Engineering Science .2019; 233(23): 7669 – 7686
[16]Wu P, Lai YC, Widelitz R, Chuong CM. Comprehensive molecular and cellular studies suggest avian scutate scales are secondarily derived from feathers, and more distant from reptilian scales. Sci Rep. 2018;8(1):16766.
[17] Cooper RL, Milinkovitch MC. Transient agonism of the sonic hedgehog pathway triggers a permanent transition of skin appendage fate in the chicken embryo. Sci Adv. 2023 May 19;9(20):eadg9619.





 扫码下载APP
扫码下载APP
 科普中国APP
科普中国APP
 科普中国
科普中国
 科普中国
科普中国